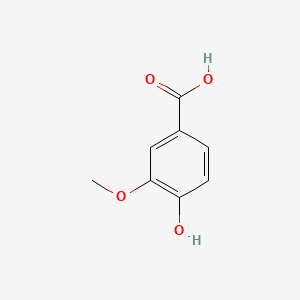
| The Content Variation of Natural Product Induced by Different Factor(s) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species Name: Amaranthus tricolor genotype VA13 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: NaCl Treatment | [1] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
On the basis of previous studies, an antioxidant enriched high yield potential genotype (Accession VA13) was selected for this investigation. This genotype was grown in pots of a rain shelter open field of Bangabandhu Sheikh Mujibur Rahman Agricultural University, Bangladesh (AEZ-28, 24° 23′ north latitude, 90° 08′ east longitude, 8.4 m.s.l.). The seeds were sown in plastic pots (15 cm in height and 40 cm length and 30 cm width) in a randomized complete block design (RCBD) with three replications. N: P2O5:K2O were applied @92:48:60 kg/ha as a split dose. First, in pot soil, @46:48:60 kg ha 1 N: P2O5:K2O and second, at 7 days after sowing (DAS) @46:0:0 kg/ha N: P2O5:K2O. The genotype was grouped into three sets and subjected to four salinity stress treatments that are, 100 mM NaCl, 50 mM NaCl, 25 mM NaCl, and control or no saline water (NS). Pots were well irrigated with fresh water every day up to 10 days after sowing (DAS) of seeds for proper establishment and vigorous growth of seedlings. Imposition of salinity stress treatment was started at 11 DAS and continued up to 40 DAS (edible stage). Saline water (100 mM NaCl, 50 mM NaCl and 25 mM NaCl) and fresh water were applied to respective pots once a day. At 40 DAS the leaves of Amaranthus tricolor were harvested. All the parameters were measured in six samples.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
At Moderate salinity stress (MSS) and Severe salinity stress (SSS) conditions, leaf color parameters and pigments, vitamins, phenolic acids, flavonoids and antioxidant capacity of A. tricolor leaves were very high compared to control condition. Hence, salt-stressed A. tricolor leaves had a good source of natural antioxidants compared to plant grown in normal cultivation practices.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
No saline water (Control)
|
Leaves | Bangabandhu |
NP Content: 9.40 ± 0.12 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
25 mM NaCl (Low salinity stress)
|
Leaves | Bangabandhu |
NP Content: 9.37 ± 0.09 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
50 mM NaCl (Moderate salinity stress)
|
Leaves | Bangabandhu |
NP Content: 12.65 ± 0.08 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
100 mM NaCl (Severe salinity stress)
|
Leaves | Bangabandhu |
NP Content: 14.89 ± 0.22 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Clausena lansium | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: Developmental Stage Variation | [2] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Clausena lansium (Lour.) Skeels leaves of four developmental stages, namely, (i) leaf buds, (ii) young leaves, (iii) mature leaves, and (iv) old leaves, were collected from three 13-year-old trees grown in wampee resources nursery of Institute of Fruit Tree Research, Guangdong Academy of Agricultural Sciences in Guangzhou, China.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increase in bound flavonoids, quercetin, and cellular antioxidant activity was observed in bound and free fractions at different stages of leaf development. Predominantly, quercetin and ferulic acid contents were high in free and bound fractions of old leaves. In addition, phenolic components depicted highly significant positive association (p < 0.05) with antioxidant activity.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Leaf buds
|
Leaves | Guangzhou, Guangdong, China |
NP Content: 1.52 ± 0.01 mg/100g
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Young leaves
|
Leaves | Guangzhou, Guangdong, China |
NP Content: 1.55 ± 0.01 mg/100g
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Mature leaves
|
Leaves | Guangzhou, Guangdong, China |
NP Content: 1.66 ± 0.01 mg/100g
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Old leaves
|
Leaves | Guangzhou, Guangdong, China |
NP Content: 1.51 ± 0.01 mg/100g
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Aleppo (green)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [3] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Aleppo (green)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00016 ± 0.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Aleppo (green)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00055 ± 0.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Aleppo (green)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00064 ± 0.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Biondonna (green)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [3] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Biondonna (green)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00026 ± 0.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Biondonna (green)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00048 ± 0.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Biondonna (green)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00049 ± 0.00003 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Carmesi (red)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [3] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Carmesi (red)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00162 ± 1.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Carmesi (red)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00428 ± 4.00002 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Carmesi (red)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00548 ± 5.00000 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Dubagold (green)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [3] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Dubagold (green)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00061 ± 0.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Dubagold (green)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.0013 ± 1.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Dubagold (green)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00264 ± 2.00008 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Dubared (red)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [3] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Dubared (red)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00071 ± 0.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Dubared (red)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00475 ± 4.00000 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Dubared (red)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00462 ± 4.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Kiribati (green)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [3] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Kiribati (green)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00018 ± 0.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Kiribati (green)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00073 ± 0.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Kiribati (green)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.0013 ± 1.0000 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Oakly (red)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [3] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Oakly (red)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00265 ± 2.00000 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Oakly (red)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00479 ± 4.00003 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Oakly (red)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00609 ± 6.00002 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Roden (red)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [3] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Roden (red)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00081 ± 0.00003 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Roden (red)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00825 ± 8.00000 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Roden (red)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00746 ± 7.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Rosaura (red)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [3] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Rosaura (red)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00028 ± 0.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Rosaura (red)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00064 ± 0.00002 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Rosaura (red)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00128 ± 1.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Spectation (red)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [3] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Spectation (red)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00044 ± 0.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Spectation (red)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00223 ± 2.00009 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Spectation (red)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00456 ± 4.00006 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Zlatava (green)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [3] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Zlatava (green)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00029 ± 0.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Zlatava (green)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.0004 ± 0.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Zlatava (green)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00115 ± 1.00002 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Zoltan (green)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [3] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Zoltan (green)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00048 ± 0.00002 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Zoltan (green)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00228 ± 2.00009 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Zoltan (green)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00246 ± 2.00005 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Thymus daenensis | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: NaCl Treatment | [4] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
The seeds were germinated in seedling plastic tray filled up with a mixture of peat moss and coco peat (1:1). The trays were placed in a glass house at the University of Massachusetts, Amherst, USA, in September 2015. After two weeks, seedlings were transplanted into plastic pots (30 cm diameter and 30 cm height, three seedlings per pot). The pots growth media consisted of 2:1:1 ratio of sieved field soil, sand, and leaf mold, respectively. The medium in each pot was supplemented with 5.8 mg P, 3.3 mg N, and 13.8 mg K to warrant the plant growth; the electrical conductivity was 0.3 mS/cm (Beckman EC meter instrument. cedar grove, New jersey, USA); the pH was 8.04. Plants were maintained at the temperature of 18-25 ℃ ; glass house conditions were set as follows: photoperiod, 16 h; relative humidity, 60-70%; light intensity, 180 µmol m 2 s-1. The source of light was a high pressure sodium lamp in addition to the day light in order to attain 16 h of light per day. Fourty days after planting (establishment period), the uniform sized plants were treated with four different salinity levels including: 0, 30, 60, and 90 mM NaCl. The irrigation with saline solution (250 mL in each pots) was performed every two days . Total amount of NaCl per pot during experiment was 8.75 L. To avoid osmotic shock the salinity treatment started with 15 mM NaCl, and was progressively increased (every two days) to reach the maximum salinity level in each treatment. To prevent water and nutrient leaching, a plastic dish was inserted under each pot and the leached water was given back to pots.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Salinity has negative impact on plant production and stimulates several physiological and biochemical modifications in thyme species. Here, a major decline in dry matter, relative water content (RWC), photosynthetic pigment contents, K+, Ca+ as well as increase in Na+ and EL was observed. Our work showed that T. daenensis was similar to T.vulgaris in terms of tolerance to high levels of salinity stress and both species were moderately tolerant to severe salt stress. It should be noted that T. daenensis, as an Iranian endemic species, competes with commercial species like T. vulgaris as a rich source of phenolic compounds. Furthermore, the relative salt tolerance of both species could be related to the exclusion of Na+ from the vascular system in order to protect tissues from salt toxicity and also to the increase of phenolic content and radical scavenging activity. Thus, the plant behavior under salinity stress may be used to boost the production of bioactive compounds to be used on an industrial level for the manufacture of nutraceuticals, functional foods and cosmetics.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
0 mM NaCl
|
Leaves | Isfahan, Iran |
NP Content: 0.6 ± 0.1 %
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
30 mM NaCl
|
Leaves | Isfahan, Iran |
NP Content: 0.6 ± 0.08 %
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
60 mM NaCl
|
Leaves | Isfahan, Iran |
NP Content: 0.7 ± 0.07 %
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
90 mM NaCl
|
Leaves | Isfahan, Iran |
NP Content: 0.80 ± 0.1 %
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Thymus vulgaris | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: NaCl Treatment | [4] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
The seeds were germinated in seedling plastic tray filled up with a mixture of peat moss and coco peat (1:1). The trays were placed in a glass house at the University of Massachusetts, Amherst, USA, in September 2015. After two weeks, seedlings were transplanted into plastic pots (30 cm diameter and 30 cm height, three seedlings per pot). The pots growth media consisted of 2:1:1 ratio of sieved field soil, sand, and leaf mold, respectively. The medium in each pot was supplemented with 5.8 mg P, 3.3 mg N, and 13.8 mg K to warrant the plant growth; the electrical conductivity was 0.3 mS/cm (Beckman EC meter instrument. cedar grove, New jersey, USA); the pH was 8.04. Plants were maintained at the temperature of 18-25 ℃ ; glass house conditions were set as follows: photoperiod, 16 h; relative humidity, 60-70%; light intensity, 180 µmol m 2 s-1. The source of light was a high pressure sodium lamp in addition to the day light in order to attain 16 h of light per day. Fourty days after planting (establishment period), the uniform sized plants were treated with four different salinity levels including: 0, 30, 60, and 90 mM NaCl. The irrigation with saline solution (250 mL in each pots) was performed every two days . Total amount of NaCl per pot during experiment was 8.75 L. To avoid osmotic shock the salinity treatment started with 15 mM NaCl, and was progressively increased (every two days) to reach the maximum salinity level in each treatment. To prevent water and nutrient leaching, a plastic dish was inserted under each pot and the leached water was given back to pots.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Salinity has negative impact on plant production and stimulates several physiological and biochemical modifications in thyme species. Here, a major decline in dry matter, relative water content (RWC), photosynthetic pigment contents, K+, Ca+ as well as increase in Na+ and EL was observed. Our work showed that T. daenensis was similar to T.vulgaris in terms of tolerance to high levels of salinity stress and both species were moderately tolerant to severe salt stress. It should be noted that T. daenensis, as an Iranian endemic species, competes with commercial species like T. vulgaris as a rich source of phenolic compounds. Furthermore, the relative salt tolerance of both species could be related to the exclusion of Na+ from the vascular system in order to protect tissues from salt toxicity and also to the increase of phenolic content and radical scavenging activity. Thus, the plant behavior under salinity stress may be used to boost the production of bioactive compounds to be used on an industrial level for the manufacture of nutraceuticals, functional foods and cosmetics.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
0 mM NaCl
|
Leaves | Isfahan, Iran |
NP Content: 0.50 ± 0.03 %
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
30 mM NaCl
|
Leaves | Isfahan, Iran |
NP Content: 0.5 ± 0.04 %
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
60 mM NaCl
|
Leaves | Isfahan, Iran |
NP Content: 0.6 ± 0.07 %
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
90 mM NaCl
|
Leaves | Isfahan, Iran |
NP Content: 0.7 ± 0.05 %
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
