| The Content Variation of Natural Product Induced by Different Factor(s) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
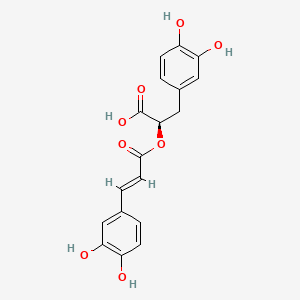
| Species Name: Dracocephalum kotschyi Boiss | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: SiO2 NPs Treatment | [1] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Seeds were treated with sulfuric acid (98%, 10 min) and then surface sterilized with 70% ethanol (v/v) for 1 min and sodium hypochlorite solution (10%, at 10 min). After sterilization, seeds were germinated on MS media (Murashige and Skoog, 1962) containing 7 g/L agar (Duchefa, Netherlands). Cultures were maintained under 16/8 h light/dark. Explants were taken from 4-week-old leaves for inoculation with bacteria strain.ATCC15834 strain of A. rhizogenes was supplied by microbial unit of the National Research Center for Genetic Engineering and Biotechnology, Tehran-Iran. Bacterial cells cultivated on LB (Luria-Bertani) culture medium (Bertani, 1952) on rotary shaker (at 26 ℃ and 180 rpm for 48 h) in the darkened state.The leaves were wounded and inoculated with bacterial suspension for 5 min and transferred to MS media containing 7 g/L agar in darkness at 25 ℃ . After 48 h treated Explants were cultured on the 1/2 MS media containing cefotaxime (500 mg/L) and indole-3-butyric acid (IBA) (2 mg/L). Hairy roots emerged at wounded sites, after 4-weeks of incubation, and then each hairy root line was isolated from explants tissue and was subcultured weekly in new media (1/2 MS hormone-free media) with appropriate antibiotic. The concentration of cefotaxime was decreased gradually and eliminated from the culture medium after 8 subcultures and axenic root cultures were obtained. Then hairy root lines were transferred to the 250 mL Erlenmeyer flasks containing 30 mL hormone- free 1/2 MS liquid medium and incubated on a rotary shaker (120 rpm) at 25 ℃ and subcultured every two week. Hairy root line, which showed sufficient growth in 1/2 MS liquid medium, was selected for further investigations.The genomic DNA was extracted from transformed hairy root lines and plant intact roots with CTAB method . Gene-specific primers from rol B were used for amplification of the 780-bp segment in PCR analysis. The primers sequences were, F:5'-ATGGATCCCAAATTGCTATTCCCCCACGA-3'and R:5'-TTAGGCTTCTTTCATTCGGTTTACTGCAGC-3'. Thirty-five PCR cycles were performed with 5 min initial denaturation at 94 ℃ , annealing steps at 60 ℃ for 80 s, extension at 72 ℃ for 90 s, and final extension step of 72 ℃ for 10 min. The amplimer were analyzed by 1% agarose gel electrophoresis.To investigate the effects of SiO2 NPs, various concentrations (0, 25, 50, 100 and 200 mg/L) of this elicitor were added to the hairy roots culture medium (1/2 MS + 3% sucrose, pH = 5.7) at the end of log phase of growth stages (21-days-old cultures). Hairy roots were incubated with elicitor for 24 and 48 h of exposure time. Hairy roots were harvested 7 days after elicitation and dried on sterile filter paper to remove excess surface moisture and were weighed before freezing by liquid nitrogen and stored at -80℃ until used to measure growth, biochemical and phytochemical analysis.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
The effect of silicon dioxide nanoparticles on production of phenolic compounds and expression rate of pal and ras genes involved in rosmarinic acid biosynthesis pathway has been investigated in D. kotschyi. SiO2 nanoparticles, used as an abiotic elicitor in our study, has appropriate optical, electrical and catalysts properties and has many applications in various industries as well as agriculture. This study clearly suggested that, in the presence of this nanoparticle, induction, production and accumulation of valuable compounds and corresponding antioxidant activity increased in hairy roots.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Mechanism |
According to the results, expression levels of the pal and ras genes were influenced by elicitor concentration and exposure time. The elicitation by SiO2 NP of 100 mg/L after 48 h of exposure time dramatically increased pal expression compared to the control. Briefly, with increasing SiO2 NP concentrations after 48 h of exposure time, the expression level of pal was also significantly induced . Similarly, ras expression was significantly raised at 48 h after treatment by increasing SiO2 NP concentration and enhanced to the greatest extent in 50 mg/L concentration. After 24 h of exposure time, the minimum level of ras expression was observed in the 200 mg/L SiO2 . Amplification products of real-time PCR were assessed with 1.8% agarose gel which was corresponded to the predicted size.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Normal condition
|
Frozen hairy roots | Iran |
NP Content: 123 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
25 mg/L SiO2 NPs + Exposure time: 24 h
|
Frozen hairy roots | Iran |
NP Content: 63.9 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
50 mg/L SiO2 NPs + Exposure time: 24 h
|
Frozen hairy roots | Iran |
NP Content: 68.8 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
100 mg/L SiO2 NPs + Exposure time: 24 h
|
Frozen hairy roots | Iran |
NP Content: 191.6 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
200 mg/L SiO2 NPs + Exposure time: 24 h
|
Frozen hairy roots | Iran |
NP Content: 19.7 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
25 mg/L SiO2 NPs + Exposure time: 48 h
|
Frozen hairy roots | Iran |
NP Content: 208.8 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
50 mg/L SiO2 NPs + Exposure time: 48 h
|
Frozen hairy roots | Iran |
NP Content: 619.7 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
100 mg/L SiO2 NPs + Exposure time: 48 h
|
Frozen hairy roots | Iran |
NP Content: 1016.6 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
200 mg/L SiO2 NPs + Exposure time: 48 h
|
Frozen hairy roots | Iran |
NP Content: 741.4 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Aleppo (green)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [2] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Aleppo (green)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00006 ± 0.00002 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Aleppo (green)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00042 ± 0.00006 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Aleppo (green)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00109 ± 1.00007 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Biondonna (green)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [2] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Biondonna (green)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00003 ± 0.00002 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Biondonna (green)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.0017 ± 1.00000 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Biondonna (green)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00301 ± 3.00005 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Carmesi (red)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [2] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Carmesi (red)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00133 ± 1.00005 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Carmesi (red)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00218 ± 2.00007 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Carmesi (red)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00141 ± 1.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Dubagold (green)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [2] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Dubagold (green)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00008 ± 0.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Dubagold (green)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.0028 ± 2.00002 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Dubagold (green)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00469 ± 4.00006 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Dubared (red)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [2] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Dubared (red)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00016 ± 0.00003 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Dubared (red)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00347 ± 3.00009 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Dubared (red)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00322 ± 3.00003 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Kiribati (green)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [2] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Kiribati (green)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00004 ± 0.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Kiribati (green)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00012 ± 0.00002 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Kiribati (green)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00167 ± 1.00006 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Oakly (red)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [2] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Oakly (red)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.0014 ± 1.00006 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Oakly (red)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00448 ± 4.00003 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Oakly (red)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00631 ± 6.00007 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Roden (red)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [2] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Roden (red)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00121 ± 1.00002 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Roden (red)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00443 ± 4.00007 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Roden (red)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00518 ± 5.00006 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Rosaura (red)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [2] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Rosaura (red)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00008 ± 0.00003 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Rosaura (red)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00091 ± 0.00008 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Rosaura (red)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00165 ± 1.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Spectation (red)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [2] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Spectation (red)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00015 ± 0.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Spectation (red)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00467 ± 4.00009 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Spectation (red)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00212 ± 2.00006 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Zlatava (green)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [2] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Zlatava (green)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00012 ± 0.00002 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Zlatava (green)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00014 ± 0.00003 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Zlatava (green)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00152 ± 1.00008 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Zoltan (green)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [2] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Zoltan (green)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00025 ± 0.00002 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Zoltan (green)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00281 ± 2.00000 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Zoltan (green)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00519 ± 5.00004 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Saponaria officinalis | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: Titanium Dioxide Nanoparticles Treatment | [3] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Different concentrations of TiO2 NPs (0, 10, 20, 30, and 50) were prepared for hairy root treatments. 0.5 g of .S. officinalis hairy roots were transferred to 250 mL Erlenmeyer flasks containing 15 mL of liquid MS culture medium with three replicates. Then, they were placed in an incubator shaker at 110 rpm and 25 ℃ in dark conditions. On the 22nd day, the liquid MS culture media containing different concentrations of nano titanium dioxide was added to Erlenmeyer flasks. 24 and 48 h after treatment, the hairy roots were taken out and transferred to the MS culture medium lacking elicitor.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
The highest rate of total phenol (9.79 mg GLA/g FW) and total flavonoid contents (1.06 mg QE/g FW) were obtained in the treated hairy roots with 50 and 30 mg/L of nano elicitor in 24 and 48 h of treatments, respectively. The maximum level of most polyphenols, such as rosmarinic acid, cinnamic acid, and rutin, was produced in 24 h of treatment. The use of TiO2 NP for 48 h with 50 mg/L concentration showed the highest production level of SO6 protein.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nano-TiO2 concentration (mg/L): 0 + Exposure time: 24h
|
hairy roots | NA |
NP Content: 47.37 mg/kg fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nano-TiO2 concentration (mg/L): 25 + Exposure time: 24h
|
hairy roots | NA |
NP Content: 57.78 mg/kg fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nano-TiO2 concentration (mg/L): 50 + Exposure time: 24h
|
hairy roots | NA |
NP Content: 58.88 mg/kg fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nano-TiO2 concentration (mg/L): 100 + Exposure time: 24h
|
hairy roots | NA |
NP Content: 58.41 mg/kg fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nano-TiO2 concentration (mg/L): 200 + Exposure time: 24h
|
hairy roots | NA |
NP Content: 64.59 mg/kg fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nano-TiO2 concentration (mg/L): 0 + Exposure time: 48h
|
hairy roots | NA |
NP Content: 47.37 mg/kg fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nano-TiO2 concentration (mg/L): 25 + Exposure time: 48h
|
hairy roots | NA |
NP Content: 57.78 mg/kg fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nano-TiO2 concentration (mg/L): 50 + Exposure time: 48h
|
hairy roots | NA |
NP Content: 44.87 mg/kg fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nano-TiO2 concentration (mg/L): 100 + Exposure time: 48h
|
hairy roots | NA |
NP Content: 62.09 mg/kg fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nano-TiO2 concentration (mg/L): 200 + Exposure time: 48h
|
hairy roots | NA |
NP Content: 30.59 mg/kg fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
