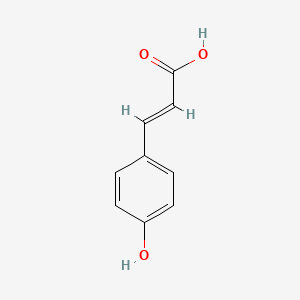
| The Content Variation of Natural Product Induced by Different Factor(s) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Species Name: Amaranthus tricolor genotype VA13 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: NaCl Treatment | [1] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
On the basis of previous studies, an antioxidant enriched high yield potential genotype (Accession VA13) was selected for this investigation. This genotype was grown in pots of a rain shelter open field of Bangabandhu Sheikh Mujibur Rahman Agricultural University, Bangladesh (AEZ-28, 24° 23′ north latitude, 90° 08′ east longitude, 8.4 m.s.l.). The seeds were sown in plastic pots (15 cm in height and 40 cm length and 30 cm width) in a randomized complete block design (RCBD) with three replications. N: P2O5:K2O were applied @92:48:60 kg/ha as a split dose. First, in pot soil, @46:48:60 kg ha 1 N: P2O5:K2O and second, at 7 days after sowing (DAS) @46:0:0 kg/ha N: P2O5:K2O. The genotype was grouped into three sets and subjected to four salinity stress treatments that are, 100 mM NaCl, 50 mM NaCl, 25 mM NaCl, and control or no saline water (NS). Pots were well irrigated with fresh water every day up to 10 days after sowing (DAS) of seeds for proper establishment and vigorous growth of seedlings. Imposition of salinity stress treatment was started at 11 DAS and continued up to 40 DAS (edible stage). Saline water (100 mM NaCl, 50 mM NaCl and 25 mM NaCl) and fresh water were applied to respective pots once a day. At 40 DAS the leaves of Amaranthus tricolor were harvested. All the parameters were measured in six samples.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
At Moderate salinity stress (MSS) and Severe salinity stress (SSS) conditions, leaf color parameters and pigments, vitamins, phenolic acids, flavonoids and antioxidant capacity of A. tricolor leaves were very high compared to control condition. Hence, salt-stressed A. tricolor leaves had a good source of natural antioxidants compared to plant grown in normal cultivation practices.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
No saline water (Control)
|
Leaves | Bangabandhu |
NP Content: 1.16 ± 0.01 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
25 mM NaCl (Low salinity stress)
|
Leaves | Bangabandhu |
NP Content: 1.25 ± 0.01 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
50 mM NaCl (Moderate salinity stress)
|
Leaves | Bangabandhu |
NP Content: 2.53 ± 0.02 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
100 mM NaCl (Severe salinity stress)
|
Leaves | Bangabandhu |
NP Content: 2.62 ± 0.03 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Brassica juncea (var. RLC-1) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: CdCl2 Treatment; Earthworms Treatment | [2] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
The experiments were conducted under controlled conditions using plastic pots having lower diameter of 7.8 cm, upper diameter of 13.5 cm and 12 cm in height. The soil was collected from the top layer (0-20 cm) from the Botanical Garden of the university. Soil was air dried crushed and sieved through 2 mm filter autoclaved at 121 ℃ for 2 h. The soil was autoclaved to exclude soil pathogens and other microorganisms if any. The autoclaved soil was poured in pots and kept in the growth chamber. The pots were filled with 500 g uncontaminated soil and partially decayed compost (cow manure) (2:1) and was used as growing medium. The cow dung was added into the soil for better performance of earthworms. A subsample of the study soil before mixing with compost was analyzed for its physicochemical characteristics. The soil used for the experiment was sandy loam soil having pH 7.8 , EC (Electrical conductivity) (µS/cm) =184.25 , TDS (Total Dissolved Solids) (mg/kg) = 130 , N (Nitrogen) (mg/kg) = 103 , P (Phosphorus) (mg/kg) = 10.6 , K (Potassium) (mg/kg) = 0.343 , %OC = 0.894, Cd (mg/kg) = ND (not detected by AAS).The Cd treatment was given by using anhydrous CdCl2 (Minimum assay: 95.0%) procured from Hi-Media laboratories. The CdCl2 anhydrous was added to the soil to make different concentrations of Cd 0.50 mM, 0.75 mM, 1.00 mM, and 1.25 mM (i.e. 56 mg/Kg , 84 mg/Kg , 112 mg/Kg and 140 mg/Kg respectively). The various treatments given are as shown below:(1)C0 (Control): (Cadmium absence);(2)C1: (0.5 mM Cd);(3)C2: (0.75 mM Cd);(4)C3: (1.00 mM Cd);(5)C4: (1.25 mM Cd).Each Cd treatment was given in soils without as well as with earthworms (WTE = without, WE = with earthworms). Earthworms (3 earthworms per pot) were inoculated after seven days of Cd treatment and incubated for 7 d in soil with earthworms. The seeds after surface sterilization were sown in soil containing different concentration of Cd and earthworms in plastic pots. These pots were kept in seed germinator under controlled conditions i.e. 25 ℃ temperature and 16:8 h dark: light photoperiod (1700 lx) for 15 d. Seedlings were harvested after 15 d followed by washing with distilled water. The growth and biochemical analysis was done on these seedlings.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased Cd uptake in plants in presence of earthworms enhances the total antioxidative capacity, metal chelating compounds and content of other antioxidants in plants grown under metal polluted soils. Earthworms can improve plant growth by improving nutrient availability to plants through their vermicasting activity. Their role in modifying soil pH and increasing metal phytoavailability made their use ideal in phytoremediation of polluted soils. Increased uptake and accumulation of Cd in plants activates the antioxidative system of plants takes place by addition of earthworms to soil.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Mechanism |
The gene expression for the key enzymes involved in organic acid metabolism was studied to understand the role of earthworms in organic acid metabolism in plants under Cd metal stress. It was observed that in comparison to control (C0) seedlings the expression of CS, SUCLG1, SDH and FH was enhanced 1.72, 1.58, 1.65 and 1.88 folds in seedlings given C4 treatment with 1.25 mM dose of Cd respectively . However, after supplementation of earthworms to Cd treated soils given C4 treatment resulted in further enhancement in expression of CS (2.53 fold), SUCLG1 (2.35 fold), SDH (2.13 fold) and FH (3.06 fold) .
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
1.00 mM CdCl2 + without earthworms
|
NA | Ludhiana, India. |
NP Content: 0.000247 ± 0.000003 mg/g
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
1.25 mM CdCl2 + without earthworms
|
NA | Ludhiana, India. |
NP Content: 0.000701 ± 0.000045 mg/g
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
0.5 mM CdCl2 + with earthworms
|
NA | Ludhiana, India. |
NP Content: 0.000552 ± 0.000076 mg/g
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Crocus sativus L. (saffron) | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: AMF Inoculation; Harvest Time Variation | [3] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
AMF Inoculation in Pot : Saffron corms with horizontal diameters of 1.3 to 2.8 cm were sown in pots (4 L; 1 corm per pot) in the last ten days of August 2016. Pots were filled with sterile quartz sand (3 L per pot) on a layer of sterilized expanded clay (1 L per pot). Corms were treated with two inocula (MycAgro Lab, Breteniere, FR), one composed of a single fungus Rhizophagus intraradices (Ri) and one of R. intraradices and Funneliformis mosseae (Ri + Fm). Ten grams of each inoculum were placed under each corm in order to guarantee the contact between the inoculum and the roots and therefore to favor the symbiosis between AMF and roots. Saffron corms used as controls were not inoculated (AMF-). Corms were not treated against fungal pathogens. A randomized block design was used with a total of 48 pots displayed in two experimental plot units (24 pots per unit) and three treatments (8 pots per treatment). Cultivation lasted for one cycle (August 2016-April 2017) in a heated glasshouse of the Department of Agricultural Forest and Food Sciences (DISAFA) of the University of Torino (Italy, 45° 06′ 23.21″ N Lat, 7° 57′ 82.8″ E Long; 293 m a.s.l.), with an average temperature of 22 ℃ during the day and 16 ℃ in the night. Irrigation water (pH 7.4, EC 505 µS cm) was added weekly (250 mL per pot) with a drip system. The corms were fertilized by fertigation (VIGORFLOR, AL.FE. srl, MN, Italy) every two weeks starting from the emergence of the spate, in quantities of 1.5 g/L of water. No flowering occurred because of the small size of the corms.AMF Inoculation in Open Field : Saffron corms with horizontal diameters of 2.5 to 3.5 cm were planted in the last ten days of August 2016 in two Alpine experimental sites located in the municipality of Morgex (45° 45′ 35″ N; 7° 02′ 37.3″ E; 1000 m a.s.l.) and Saint Cristophe (45° 45′ 06″ N; 7° 20′ 37″ E; 700 m a.s.l.) in Italy and cultivation lasted for two cycles (2016-2017 and 2017-2018). Both sites were cultivated with saffron for at least the previous three years. Before starting the experiment both fields were milled. To assess the effects of AMF inocula on saffron cultivation and production, the same treatments used in the pot trial were applied (Ri, Ri + Fm or AMF-). A randomized block design was used, with three experimental plot units (blocks). Each plot unit consisted of 56 corms, planted in a 1.44 m2 area (39 corms m-2). Inter-row planting distance was of 7 cm, while between-row distance was 25 cm. Plots were separated from each other with at least 4 m distance. Before planting, 10 g of inoculum was placed under the corms to ensure contact between plant and the treatment. Irrigation was provided when needed and hand weeding control was conducted during cultivation, while no preplanting fertilization, tillage, or treatments against pathogens were applied. The two Alpine sites were characterized by semicontinental climate, with a long and cold winter . In general, both sites had a sandy-loam texture according to the USDA classification and similar chemical characteristics.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
The inoculum composed by R. intraradices and F. mosseae was particularly effective in increasing flower production and saffron yield, while R. intraradices alone increased the content of some bioactive compounds-picrocrocin, quercitrin, crocin II-as well as antioxidant activity.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Harvesting time: 2016-2017
|
Powdered saffrons | Italy |
NP Content: 23 mg/100g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Harvesting time: 2017-2018
|
Powdered saffrons | Italy |
NP Content: 23 mg/100g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Rhizophagus intraradices and Funneliformis mosseae inoculation
|
Powdered saffrons | Italy |
NP Content: 23 mg/100g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Rhizophagus intraradices inoculation
|
Powdered saffrons | Italy |
NP Content: 23 mg/100g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Non-AMF inoculation (Control)
|
Powdered saffrons | Italy |
NP Content: 23 mg/100g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Cucumis sativus L. cv. Jincun 2 | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: Low Temperature Treatment; AMF Inoculation | [4] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
AMF inocula [Funneliformis mosseae (T.H. Nicolson & Gerd.) C. Walker & A. Schubler] consisting of spores, soil, hyphae and infected clove (Trifolium repens L.) root fragment from a stock culture of F. mosseae.The inoculated dosage was 10 g of inocula per pot containing approximately 720 spores calculated by microscopy before experiment. There was 2100 infective propagules/g in the inoculum as determined by MPN assay . Seeds were surface sterilized by immersion in 70% ethanol for 5 min, rinsed four times with distilled water, and placed on wet filter paper in Petri dishes at 28 ℃ for germination. After 3 days, the germinate seeds were transplanted into 13 cm × 13 cm plastic pots containing 0.88 kg organized soil substrate (organic manure, soil and decomposed straw = 1:2:1). Half of the pots (AM plants) were inoculated with 10 g of F. mosseae per pot. Non-AM plants received the same weight of autoclaved inocula. The inocula were placed adjacent to each seeding root. The organic substrate was collected from the greenhouse of Institute of Vegetables and Flowers, CAAS and sterilized for 4 h at 160 ℃ , with the chemical properties as follows: pH 7.26, 11.1% organic matter, 150 mg/kg available phosphorus, 451 mg/kg available nitrogen and 518 mg/kg available potassium. The experimental pots were placed in solar greenhouse at an average temperature of 28 ℃ /20 ℃ (day/night) with photon flux density of 600 µmol m-2 s-1 and 85% relative humidity.The seedlings uniformed in size were transferred to a growth chamber subjected to different temperature conditions at 20 days after inoculation. The experimental design consisted of four treatments crossing two mycorrhizal inoculations levels (non-AMF and F. mosseae) with two temperature levels with photon flux density of 100 µmol m-2 s-1 (25 ℃ /15 ℃ , 15 ℃ /10 ℃ , day/night). (1) Normal temperature (NT): 10 g of sterilized inoculua, 25 ℃ /15 ℃ (day/night); (2) AMF-inoculation (AMF): 10 g of inocula, 25 ℃ /15 ℃ (day/night); (3) low temperature (LT): 10 g of sterilized inocula, 15 ℃ /10 ℃ (day/night); (4) AMF-inoculation under low temperature (AMF + LT): 10 g of inocula, 15 ℃ /10 ℃ (day/night).The experimental design was a completely randomized block design and thirty plants were arranged in each replication. On 45 days after inoculation, phenolic compounds contents, enzymes and gene transcription were determined.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
AMF-inoculated cucumber seedlings had significant higher fresh weight and dry weight than non-AMF inoculated control plants under both normal (25/15 ℃ ) and low temperature (15/10 ℃ ) treatment. Under chilling stress, AMF inoculation significantly improved the content of related secondary metabolites including phenols, flavonoids, lignin, DPPH activity and phenolic compounds compared with the non-AMF control. Furthermore, large increments were observed in a number of enzymatic activities related to secondary metabolism and antioxidant system in AMF-inoculated seedlings under low temperature, such as glucose-6-phosphate dehydrogenase (G6PDH), shikimate dehydrogenase (SKDH), phenylalanine ammonia-lyase (PAL), cinnamyl alcohol dehydrogenase (CAD), polyphenol oxidase (PPO), guaiacol peroxidase (G-POD), caffeic acid peroxidase (CA-POD) and chlorogenic acid peroxidase (CGA-POD). As well, the expression of stress-related marker genes was enhanced in AMF-inoculated seedlings in comparison with the non-AMF control. Furthermore, AMF symbiosis decreased hydrogen peroxide (H2O2) content under low temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Mechanism |
Consistent with the changes of enzyme activities, the relative transcriptional level of genes related to secondary metabolism increased significantly in response to AM fungi inoculation in cucumber roots . Moreover, under chilling stress, the expression levels of WRKY30, PR-1, C4H, CCOMT, CAD, G6PDH, PAL, LPO, and POD genes in the AMF-inoculated seedling were 2.46, 2.90, 1.84, 2.47, 1.96, 2.52, 1.89 and 1.93 folds respectively compared with those under low temperature alone.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Normal temperature: 10 g of sterilized inoculua (Funneliformis mosseae), 25/15 ℃ (day/night)
|
Leaves | NA |
NP Content: 1.45 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Normal temperature: 10 g of sterilized inoculua (Funneliformis mosseae), 25/15 ℃ (day/night)
|
Leaves | NA |
NP Content: 18.45 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
AMF-inoculation: 10 g of inocula (F. mosseae), 25/15 ℃ (day/night)
|
Leaves | NA |
NP Content: 2.42 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
AMF-inoculation: 10 g of inocula (F. mosseae), 25/15 ℃ (day/night)
|
Leaves | NA |
NP Content: 34.47 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Low temperature: 10 g of sterilized inocula (F. mosseae), 15/10 ℃ (day/night)
|
Leaves | NA |
NP Content: 2.26 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Low temperature: 10 g of sterilized inocula (F. mosseae), 15/10 ℃ (day/night)
|
Leaves | NA |
NP Content: 29.47 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
AMF-inoculation under low temperature: 10 g of inocula (F. mosseae), 15/10 ℃ (day/night)
|
Leaves | NA |
NP Content: 2.83 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
AMF-inoculation under low temperature: 10 g of inocula (F. mosseae), 15/10 ℃ (day/night)
|
Leaves | NA |
NP Content: 52.38 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Dracocephalum kotschyi Boiss | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: SiO2 NPs Treatment | [5] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Seeds were treated with sulfuric acid (98%, 10 min) and then surface sterilized with 70% ethanol (v/v) for 1 min and sodium hypochlorite solution (10%, at 10 min). After sterilization, seeds were germinated on MS media (Murashige and Skoog, 1962) containing 7 g/L agar (Duchefa, Netherlands). Cultures were maintained under 16/8 h light/dark. Explants were taken from 4-week-old leaves for inoculation with bacteria strain.ATCC15834 strain of A. rhizogenes was supplied by microbial unit of the National Research Center for Genetic Engineering and Biotechnology, Tehran-Iran. Bacterial cells cultivated on LB (Luria-Bertani) culture medium (Bertani, 1952) on rotary shaker (at 26 ℃ and 180 rpm for 48 h) in the darkened state.The leaves were wounded and inoculated with bacterial suspension for 5 min and transferred to MS media containing 7 g/L agar in darkness at 25 ℃ . After 48 h treated Explants were cultured on the 1/2 MS media containing cefotaxime (500 mg/L) and indole-3-butyric acid (IBA) (2 mg/L). Hairy roots emerged at wounded sites, after 4-weeks of incubation, and then each hairy root line was isolated from explants tissue and was subcultured weekly in new media (1/2 MS hormone-free media) with appropriate antibiotic. The concentration of cefotaxime was decreased gradually and eliminated from the culture medium after 8 subcultures and axenic root cultures were obtained. Then hairy root lines were transferred to the 250 mL Erlenmeyer flasks containing 30 mL hormone- free 1/2 MS liquid medium and incubated on a rotary shaker (120 rpm) at 25 ℃ and subcultured every two week. Hairy root line, which showed sufficient growth in 1/2 MS liquid medium, was selected for further investigations.The genomic DNA was extracted from transformed hairy root lines and plant intact roots with CTAB method . Gene-specific primers from rol B were used for amplification of the 780-bp segment in PCR analysis. The primers sequences were, F:5'-ATGGATCCCAAATTGCTATTCCCCCACGA-3'and R:5'-TTAGGCTTCTTTCATTCGGTTTACTGCAGC-3'. Thirty-five PCR cycles were performed with 5 min initial denaturation at 94 ℃ , annealing steps at 60 ℃ for 80 s, extension at 72 ℃ for 90 s, and final extension step of 72 ℃ for 10 min. The amplimer were analyzed by 1% agarose gel electrophoresis.To investigate the effects of SiO2 NPs, various concentrations (0, 25, 50, 100 and 200 mg/L) of this elicitor were added to the hairy roots culture medium (1/2 MS + 3% sucrose, pH = 5.7) at the end of log phase of growth stages (21-days-old cultures). Hairy roots were incubated with elicitor for 24 and 48 h of exposure time. Hairy roots were harvested 7 days after elicitation and dried on sterile filter paper to remove excess surface moisture and were weighed before freezing by liquid nitrogen and stored at -80℃ until used to measure growth, biochemical and phytochemical analysis.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
The effect of silicon dioxide nanoparticles on production of phenolic compounds and expression rate of pal and ras genes involved in rosmarinic acid biosynthesis pathway has been investigated in D. kotschyi. SiO2 nanoparticles, used as an abiotic elicitor in our study, has appropriate optical, electrical and catalysts properties and has many applications in various industries as well as agriculture. This study clearly suggested that, in the presence of this nanoparticle, induction, production and accumulation of valuable compounds and corresponding antioxidant activity increased in hairy roots.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Mechanism |
According to the results, expression levels of the pal and ras genes were influenced by elicitor concentration and exposure time. The elicitation by SiO2 NP of 100 mg/L after 48 h of exposure time dramatically increased pal expression compared to the control. Briefly, with increasing SiO2 NP concentrations after 48 h of exposure time, the expression level of pal was also significantly induced . Similarly, ras expression was significantly raised at 48 h after treatment by increasing SiO2 NP concentration and enhanced to the greatest extent in 50 mg/L concentration. After 24 h of exposure time, the minimum level of ras expression was observed in the 200 mg/L SiO2 . Amplification products of real-time PCR were assessed with 1.8% agarose gel which was corresponded to the predicted size.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Normal condition
|
Frozen hairy roots | Iran |
NP Content: 12.06 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
25 mg/L SiO2 NPs + Exposure time: 24 h
|
Frozen hairy roots | Iran |
NP Content: 2.58 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
50 mg/L SiO2 NPs + Exposure time: 24 h
|
Frozen hairy roots | Iran |
NP Content: 5.86 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
100 mg/L SiO2 NPs + Exposure time: 24 h
|
Frozen hairy roots | Iran |
NP Content: 9.95 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
200 mg/L SiO2 NPs + Exposure time: 24 h
|
Frozen hairy roots | Iran |
NP Content: 3.14 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
25 mg/L SiO2 NPs + Exposure time: 48 h
|
Frozen hairy roots | Iran |
NP Content: 2.36 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
50 mg/L SiO2 NPs + Exposure time: 48 h
|
Frozen hairy roots | Iran |
NP Content: 4.56 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
100 mg/L SiO2 NPs + Exposure time: 48 h
|
Frozen hairy roots | Iran |
NP Content: 8.11 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
200 mg/L SiO2 NPs + Exposure time: 48 h
|
Frozen hairy roots | Iran |
NP Content: 17.63 µg/g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Fragaria × ananassa Duch. | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: Nitrogen Treatment; AMF Inoculation | [6] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
The experiment was conducted in a 'shade'-type greenhouse with 30% shade at the Instituto de Investigaciones Agropecuarias y Forestales (IIAF), Universidad Michoacana de San Nicolas de Hidalgo (UMSNH), Morelia, Michoacan, Mexico. Maximum and minimum temperatures in the greenhouse varied between 28 and 32 ℃ and between 8 and 18 ℃ respectively. Plants of the strawberry cultivar 'Aromas' were used that had previously been grown in a sterilised (95 ℃ water/steam, 40 min) substrate of coconut fibre/perlite (1:3 v/v) under greenhouse conditions. Before the experiment was established, the absence of AMF in the roots was verified by the ink and vinegar technique, modifying the duration of immersion in KOH and ink/vinegar solution (7 and 5 min respectively). Before planting, roots were disinfected by submerging them for 20 s in 20 g/L sodium hypochlorite solution and rinsing them in water. The inoculum was prepared with spores of Glomus intraradices cultivated in liquid medium (3.5 × 106 spores/L, 90% viability; Premier Tech Biotechnologies Company, Quebec, Canada), which was diluted with fitagel (Sigma P-8169, Saint Louis, MO, USA) solution at 50 g/L to obtain a final concentration of about 5 × 104 spores/L. The viability of spores was determined according to the method of An and Hendrix. Eighteen days after setting up the experiment, each plant received 2 mL of inoculum applied directly to the recently formed roots. One month later, after staining, the percentage of root colonisation was determined by the gridline intersect method. The experiment was organised as a full factorial, completely randomised design with two factors: inoculation (two levels: mycorrhizal and non-mycorrhizal plants) and N concentration in the nutrient solution (three levels: 3, 6 and 18 mmol/L). The six treatments were replicated four times, producing 24 experimental units with ten plants each. Every second day, all plants were irrigated up to substrate saturation. Nitrogen was supplied as NO and the cation/anion ratio was kept constant by varying the concentration of SO. When N was below 18 mmol/L, the cation concentrations were maintained as follows: K+, 3; Ca2+, 3.5; Mg2+, 1.5 mmol/L. They were increased in the 18 mmol/L N treatment: K+, 6.5; Ca2+, 7.5; Mg2+, 3.25 mmol/L. In all nutrient solutions the concentration of phosphorus (P) was 0.3 mmol/L. The other nutrients in the solutions were: H3BO3, 20; CuSO4. 5H2O, 0.5; Fe-EDTA (Ethylenediaminetetraacetic acid iron (III) sodium salt), 15; MnSO4.H2O, 12; (NH4)6Mo7O24 . 4H2O, 0.05; ZnSO4 . 7H2O, 3 µmol/L. The pH was adjusted to 5.5 at every application date.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Mycorrhization did not modify the weight, diameter or length of strawberry fruits but had a negative effect on most colour parameters. Moreover, fruits of mycorrhizal plants had higher K and Cu concentrations and showed greater accumulation of most phenolic compounds.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nitrogen concentration (mmol/L): 3
|
Mature fruits | Morelia, Michoacan, Mexico |
NP Content: 0.54163 g/kg dry matter
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nitrogen concentration (mmol/L): 6
|
Mature fruits | Morelia, Michoacan, Mexico |
NP Content: 0.50332 g/kg dry matter
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nitrogen concentration (mmol/L): 18
|
Mature fruits | Morelia, Michoacan, Mexico |
NP Content: 0.68204 g/kg dry matter
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Glomus intraradices inoculation
|
Mature fruits | Morelia, Michoacan, Mexico |
NP Content: 0.57131 g/kg dry matter
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Non-AMF inoculation
|
Mature fruits | Morelia, Michoacan, Mexico |
NP Content: 0.58001 g/kg dry matter
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nitrogen concentration (mmol/L): 3 + G. intraradices inoculation
|
Mature fruits | Morelia, Michoacan, Mexico |
NP Content: 0.59038 g/kg dry matter
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nitrogen concentration (mmol/L): 3 + Non-AMF inoculation
|
Mature fruits | Morelia, Michoacan, Mexico |
NP Content: 0.49287 g/kg dry matter
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nitrogen concentration (mmol/L): 6 + G. intraradices inoculation
|
Mature fruits | Morelia, Michoacan, Mexico |
NP Content: 0.50674 g/kg dry matter
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nitrogen concentration (mmol/L): 6 + Non-AMF inoculation
|
Mature fruits | Morelia, Michoacan, Mexico |
NP Content: 0.49989 g/kg dry matter
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nitrogen concentration (mmol/L): 18 + G. intraradices inoculation
|
Mature fruits | Morelia, Michoacan, Mexico |
NP Content: 0.61679 g/kg dry matter
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nitrogen concentration (mmol/L): 18 + Non-AMF inoculation
|
Mature fruits | Morelia, Michoacan, Mexico |
NP Content: 0.74729 g/kg dry matter
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Aleppo (green)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [7] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Aleppo (green)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00533 ± 5.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Aleppo (green)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00592 ± 5.00006 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Aleppo (green)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00529 ± 5.00003 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Biondonna (green)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [7] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Biondonna (green)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00562 ± 5.00004 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Biondonna (green)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00579 ± 5.00004 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Biondonna (green)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00595 ± 5.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Carmesi (red)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [7] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Carmesi (red)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00608 ± 6.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Carmesi (red)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.0058 ± 5.0000 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Carmesi (red)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00676 ± 6.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Dubagold (green)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [7] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Dubagold (green)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.0051 ± 5.00007 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Dubagold (green)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00647 ± 6.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Dubagold (green)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00685 ± 6.00005 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Dubared (red)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [7] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Dubared (red)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00584 ± 5.00006 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Dubared (red)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00639 ± 6.00007 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Dubared (red)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00653 ± 6.00002 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Kiribati (green)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [7] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Kiribati (green)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00627 ± 6.00000 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Kiribati (green)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00592 ± 5.00004 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Kiribati (green)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00664 ± 6.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Oakly (red)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [7] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Oakly (red)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00573 ± 5.00002 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Oakly (red)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00597 ± 5.00002 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Oakly (red)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00629 ± 6.00006 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Roden (red)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [7] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Roden (red)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00577 ± 5.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Roden (red)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00578 ± 5.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Roden (red)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00613 ± 6.00006 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Rosaura (red)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [7] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Rosaura (red)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00551 ± 5.00004 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Rosaura (red)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00579 ± 5.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Rosaura (red)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00592 ± 5.00009 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Spectation (red)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [7] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Spectation (red)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00581 ± 5.00007 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Spectation (red)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00663 ± 6.00004 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Spectation (red)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00772 ± 7.00003 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Zlatava (green)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [7] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Zlatava (green)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00573 ± 5.00003 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Zlatava (green)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.0057 ± 5.00001 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Zlatava (green)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00572 ± 5.00002 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lactuca sativa var. crispa 'Zoltan (green)' | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: UV Radiation Treatment; Cultivar Comparison | [7] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Twelve cultivars of leaf-type lettuce (Lactuca sativa var. crispa) were selected for the study. This type of lettuce forms open heads with loose leaves that do not close to cover younger leaves. Six green-colored cultivars originated from Semo, a.s., Czech Republic (Dubagold, Zlatava, and Zoltan) and Bejo Zaden B.V., Netherlands (Aleppo, Biondonna, and Kiribati); six red-colored cultivars were also from Semo (Dubared, Roden, and Rosaura) and Bejo Zaden (Carmesi, Oakly, and Spectation). The experiments were performed in the spring period (April, May). Lettuce seeds were sown in plastic pots and germinated under standard laboratory conditions (ca. 21 ℃ , 12-hour photoperiod). After germination, the lettuce plants were transplanted into a growth chamber (air-conditioned box model MC1750 (Snijders Scientific, Tilburg, Netherlands) and grown under 14/10 h (day/night) photoperiod, 21/18 ℃ temperature, 60% humidity, and 250 µmol.m -2.s-1 light intensity. The commercial peat substrate (Klasmann, Germany) was used (pH 6.0, nutrient content: N: 220 mg/L, P2O5: 110 mg/L, K2O: 220 mg/L, Mg: 80 mg/L). Approximately at the stage of second, fully expanded, true leaf the plants were transplanted to 0.5 l pots and kept at the same conditions for seven days to recover. After recovery, the plants were transferred into one of the three experimental conditions described below. Plants were watered regularly to avoid drought stress. Considering a high level of nutrients in the substrate and a short duration of the experiments, no additional nutrition was applied to plants.The experiments were held at SAU in Nitra (48° 19′ 7″ N, 18° 4′ 55″ E, 144 m asl). To distinguish the effects of UV radiation from other environmental factors such as temperature, humidity, and light intensity, the plants were grown in three different environments: 1. direct sunlight (outdoor conditions with high UV), 2. under clear glass (outdoor conditions with low UV), and 3. greenhouse (indoor conditions with low UV).Plants grown under direct sunlight conditions were placed into a vegetation cage (a walk-in cage surrounded by the thin wire mesh from the top and side to protect the experimental plants against birds and animals) and exposed to almost unrestricted sunlight and ambient temperature and humidity. Plants were watered as needed to achieve a fully hydrated state. Temperature outdoors was monitored.Plants grown under clear glass were placed in similar environmental conditions as those cultivated under direct sunlight, but were grown in the glass shelter constructed from clear glass sheets (thickness of 8 mm). The clear glass sheets were positioned such as to eliminate UV light coming to plants from the south and above. The backside (oriented to the north) of this glass shelter was covered by the plastic-coated wire mesh not impeding the flow of air, so the temperature and other conditions were almost identical to fully open outdoor conditions. Temperature outdoors and under the glass sheets was occasionally compared using hand-held thermometers, showing only insignificant differences. The glass cover lowered the intensity of photosynthetically active radiation (PAR) by 10-15% at noon due to absorbance and reflectance of radiation by the glass. The overhang of the glass shelter, wire mesh from the north part as well as composition of buildings from the north west and east direction was very favorable to prevent excessive access of diffuse UV radiation. Thus, despite UV radiation is not fully eliminated, its level represents only a small fraction compared to the direct UV radiation incident to plants exposed to direct sunlight outdoors.Plants grown indoors were placed in a regular greenhouse constructed from clear glass that eliminated approximately 15-20% of PAR intensity at noon. Light intensity in the greenhouse reached almost 1,000 µmol photons m-2 s-1 during sunny days, therefore it can still be regarded as fully saturating or excessive radiation, similar to that at outdoor conditions. Temperature in the greenhouse was lowered during the day by the automated ventilation, but air vents were closed during the night. Temperature in the greenhouse was substantially higher than outdoors (environmental conditions 1 and 2). During the experiment, the night temperature in the greenhouse ranged between 15 and 20 ℃ , whereas the daily maximum temperature oscillated mostly between 20 and 32 ℃ . The maximum temperature of 35 ℃ was reached during a few of the warmest days. In each environment, plants were grown in the randomized complete block design, with weekly rotations of plant positions. Four healthy, well-developed plants from each cultivar were selected for analyses from each of the three environments. Non-destructive analyses started 30 days after sowing and continued for another 30 days. The complete above-ground parts of the plants were harvested at the end of the experiment (60 days after sewing) and were used for destructive analyses.Comparison between environments is based on the assumption that the plants grown under the glass sheets outdoors were exposed to similar light and low UV conditions as the plants in the greenhouse, but the temperature conditions were similar to those in direct sunlight outdoors. By comparing accumulation of phenolic compounds in plants grown in the three environments, we could distinguish the effects caused by UV radiation from those caused by the temperature.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Increased accumulation of total phenolics, flavonoids, anthocyanins, and phenolic acids was observed in direct sunlight conditions outdoors as compared to the greenhouse conditions with low UV radiation, but elevated day and night temperatures. The level of UV radiation played a dominant role in the accumulation of flavonoids, anthocyanins and methoxycinnamic acid; while temperature was a major factor affecting concentrations of phenolic acids, mostly rosmarinic, p-anisic and vanillic acid. The concentrations of compounds estimated with the non-invasive fluorescence excitation ratio method were highly consistent with those obtained by standard analytical approaches.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Zoltan (green)': Greenhouse (indoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00584 ± 5.00005 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Zoltan (green)': Under clear glass (outdoor conditions with low UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00648 ± 6.00003 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Lactuca sativa var. crispa 'Zoltan (green)': Direct sunlight (outdoor conditions with high UV)
|
Leaves | Nitra, Slovak |
NP Content: 0.00652 ± 6.00003 mg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Lentil var. Tina | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: H2O2 Treatment; Mannitol Treatment; NaCl Treatment; High Temperature Treatment; Low Temperature Treatment | [8] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Seeds were sterilized in 1% (v/v) sodium hypochloride (Sigma-Aldrich, USA) for 10 min, then drained and washed with distilled water until they reached neutral pH. They were placed in distilled water and soaked for 6 h at 25 ℃ . Seeds were dark germinated for 8 days in a growth chamber (SANYO MLR-350H) on Petri dishes (125 mm) lined with absorbent paper. Seedlings were watered with 5 ml of Milli-Q water daily. Sprout (8-day-old) samples were gently collected, weighed (fresh mass), rapidly frozen and kept in polyethylene bags at -20 ℃ . For each treatment, three replicates were performed.Elicitation conditions were selected in previous screening studies. For the experiments, temperature (4 ℃ and 40 ℃ - TC and TH, respectively), H2O2 (20 mM and 200 mM - Ox1 and Ox2, respectively), mannitol (200 mM and 600 mM - Os1 and Os2, respectively) and NaCl (100 mM and 300 mM - S-Os1 and S-Os2, respectively) were selected as abiotic elicitors. All solutions were freshly prepared before each application. Mannitol (Os1, Os2), NaCl (S-O1, S-O2) and H2O2 (Ox1) treatments were applied by watering daily (not soaking) 2-day-old sprouts with 5 ml of test solution. For Ox2 (200 mM H2O2) treatment 2-day-old seedlings were only once watered with 5 ml of 200 mM H2O2 and then cultivated under standard conditions. For temperature conditioning treatment, 2-day-old sprouts were incubated at 4 ℃ and 40 ℃ (TC and TH, respectively) for 1 h and then cultivated under standard conditions. Sprout (8-day-old) samples were gently collected, weighed (fresh mass), rapidly frozen and kept in polyethylene bags at -20 ℃ .
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Application of abiotic elicitors (environmental shocks) was an effective method for improvement of sprout pro-health potential via an increase of phenolic contents and subsequent elevation of antioxidant potential. Innovative application of elicitors on 2-day-old sprouts (not seed) allowed the elimination of the unfavorable influence of the factors employed on germination yield and biomass production. Assuming that the optimal germination conditions are those which most effectively increase the antioxidant potential without any negative influence on biomass accumulation and nutritional quality the elicitation with 20 mM H2O2 for the future applications is recommended.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Normal condition
|
Sprouts | NA |
NP Content: 0.03308 ± 0.001 mg/g flour
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Induction with 20 mM H2O2
|
Sprouts | NA |
NP Content: 0.05143 ± 0.0027 mg/g flour
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Induction with 200 mM H2O2
|
Sprouts | NA |
NP Content: 0.04212 ± 0.0029 mg/g flour
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Induction with 200 mM mannitol
|
Sprouts | NA |
NP Content: 0.03752 ± 0.001 mg/g flour
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Induction with 600 mM mannitol
|
Sprouts | NA |
NP Content: 0.03411 ± 0.0024 mg/g flour
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Induction with 100 mM NaCl
|
Sprouts | NA |
NP Content: 0.03095 ± 0.0015 mg/g flour
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Induction with 300 mM NaCl
|
Sprouts | NA |
NP Content: 0.02921 ± 0.0026 mg/g flour
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Induction at 4 ℃
|
Sprouts | NA |
NP Content: 0.02485 ± 0.0009 mg/g flour
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Induction at 40 ℃
|
Sprouts | NA |
NP Content: 0.03502 ± 0.0063 mg/g flour
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Rubus idaeus | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: Cultivar Comparison; Organic Fertilization; Traditional Fertilization | [9] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
The experiment was carried out in 2013. Leaves of five raspberry cultivars ('Polana', 'Polka', 'Tulameen', 'Laszka' and 'Glen Ample') were collected at the time of cultivation. Three organic and neighborhood conventional farms were used for experimental purposes. From one cultivar (one field plot), 3-4 plants were chosen, which were analyzed separately. One sample consisted of 10 leaves. The farm was treated as a replication. [organic farm no. 1 Localization: akroczym(52° 26″ N 20° 36″ E), Type of Soil: sandy middle soil IVa and IVb category (15% floatable particles) pH 5.5, Kind of Fertilizer: cow manure, Dose of Fertilizers and Time of Given: 35 t/ha one year before raspberry planting, Plant Protection System: Grevit 200 SL; organic farm no. 2 Localization: Zaluski (52° 37″ N 20° 22″ E), Type of Soil: sandy middle soil, sandy-clay IV category (20% floatable particles), pH 5.5, Kind of Fertilizer:cow manure, Dose of Fertilizers and Time of Given: 30 t/ha one year before raspberry planting, Plant Protection System: no protection; organic farm no. 3 Localization: Radzanow(51° 33″ N 20° 51″ E), Type of Soil: sandy middle soil IVa and III category (10% floatable particles), pH 6.0, Kind of Fertilizer:sheep manure, green manure, Dose of Fertilizers and Time of Given: 10 t/ha and 15 t/ha one year before raspberry planting, Plant Protection System: Bioczos 33 SL, Grevit 200 SL; conventional farm no. 1 Localization: Czerwinsk nad Wisla (52° 23″ N 20° 20″ E), Type of Soil: sandy-loamy middle soil IV and III category (20% floatable particles), pH 5.5, Kind of Fertilizer: Hydrocomplex 12-11-18; Superba 8-11-36, Dose of Fertilizers and Time of Given: (200 kg/ha, 150 kg/ha) in autumn a year before raspberry planting; 3 doses in time of cultivation, Plant Protection System: Signum 33 WG, Miros 20 SP; conventional farm no. 2 Localization: Czerwinsk nad Wisla (52° 23″ N 20° 20″ E), Type of Soil: sandy-loamy middle soil IV and III category (25% floatable particles), pH 5.5, Kind of Fertilizer: amonium nitrate, polyphosphate, magnesium sulphate, Dose of Fertilizers and Time of Given: in autumn a year before raspberry planting; 3 doses in time of cultivation, Plant Protection System: Calypso 480 SC, Miros 20 SP, Zato 50 WG; conventional farm no. 3 Localization: Czerwinsk nad Wisla(52° 25″ N 20° 23″ E), Type of Soil: sandy-clay middle soil II and III category (20% floatable particles) pH 6.0, Kind of Fertilizer:Rosafert 5-12-24-3, Dose of Fertilizers and Time of Given: 250 kg/ha in autumn a year before raspberry planting; 4 doses in time of cultivation, Plant Protection System: Calypso 480 SC, Miros 20 SP, Zato 50 WG].
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
Compared with conventional raspberry leaves, organic raspberry leaves were characterized by a significantly higher content of dry matter, total polyphenols, total phenolic acids, chlorogenic acid, caffeic acid, salicylic acid and quercetin-3-O-rutinoside; moreover, the organic leaves were characterized by higher antioxidant activity. Among examined cultivars, 'Polka' c. was characterized by the highest antioxidant status. However, raspberry leaves from conventional farms contained more total carotenoids, violaxanthin, alpha-carotene, beta-carotene, total chlorophyll and individual forms of chlorophylls: a and b.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Cultivation System: organic farm
|
Leaves | Poland |
NP Content: 14.77 ± 0.83 mg/100g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Cultivation System: conventional farm
|
Leaves | Poland |
NP Content: 25.02 ± 3.89 mg/100g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Rubus idaeus cv. Polana
|
Leaves | Poland |
NP Content: 28.81 ± 6.15 mg/100g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Rubus idaeus cv. Polka
|
Leaves | Poland |
NP Content: 22.67 ± 5.92 mg/100g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Rubus idaeus cv. Tulameen
|
Leaves | Poland |
NP Content: 13.92 ± 1.17 mg/100g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Rubus idaeus cv. Laszka
|
Leaves | Poland |
NP Content: 10.10 ± 2.43 mg/100g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Rubus idaeus cv. Glen Ample
|
Leaves | Poland |
NP Content: 25.14 ± 2.24 mg/100g fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Saponaria officinalis | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: Titanium Dioxide Nanoparticles Treatment | [10] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
Different concentrations of TiO2 NPs (0, 10, 20, 30, and 50) were prepared for hairy root treatments. 0.5 g of .S. officinalis hairy roots were transferred to 250 mL Erlenmeyer flasks containing 15 mL of liquid MS culture medium with three replicates. Then, they were placed in an incubator shaker at 110 rpm and 25 ℃ in dark conditions. On the 22nd day, the liquid MS culture media containing different concentrations of nano titanium dioxide was added to Erlenmeyer flasks. 24 and 48 h after treatment, the hairy roots were taken out and transferred to the MS culture medium lacking elicitor.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
The highest rate of total phenol (9.79 mg GLA/g FW) and total flavonoid contents (1.06 mg QE/g FW) were obtained in the treated hairy roots with 50 and 30 mg/L of nano elicitor in 24 and 48 h of treatments, respectively. The maximum level of most polyphenols, such as rosmarinic acid, cinnamic acid, and rutin, was produced in 24 h of treatment. The use of TiO2 NP for 48 h with 50 mg/L concentration showed the highest production level of SO6 protein.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nano-TiO2 concentration (mg/L): 0 + Exposure time: 24h
|
hairy roots | NA |
NP Content: 35.89 mg/kg fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nano-TiO2 concentration (mg/L): 25 + Exposure time: 24h
|
hairy roots | NA |
NP Content: 32.65 mg/kg fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nano-TiO2 concentration (mg/L): 50 + Exposure time: 24h
|
hairy roots | NA |
NP Content: 34.22 mg/kg fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nano-TiO2 concentration (mg/L): 100 + Exposure time: 24h
|
hairy roots | NA |
NP Content: 40.77 mg/kg fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nano-TiO2 concentration (mg/L): 200 + Exposure time: 24h
|
hairy roots | NA |
NP Content: 44.4 mg/kg fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nano-TiO2 concentration (mg/L): 0 + Exposure time: 48h
|
hairy roots | NA |
NP Content: 35.89 mg/kg fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nano-TiO2 concentration (mg/L): 25 + Exposure time: 48h
|
hairy roots | NA |
NP Content: 32.65 mg/kg fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nano-TiO2 concentration (mg/L): 50 + Exposure time: 48h
|
hairy roots | NA |
NP Content: 40.09 mg/kg fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nano-TiO2 concentration (mg/L): 100 + Exposure time: 48h
|
hairy roots | NA |
NP Content: 53.34 mg/kg fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Nano-TiO2 concentration (mg/L): 200 + Exposure time: 48h
|
hairy roots | NA |
NP Content: 53.68 mg/kg fresh weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Species Name: Vitis vinifera cv. Pinot noir | |||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Name: Drought Stress Treatment | [11] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Experiment Detail |
3-year old single shoot V. vinifera plants (cultivar Pinot noir 18 Gm grafted on Kober 5BB, 51 plants) potted in 3L pots in a sandy loam soil were used. All plants were well watered (200 mL per day) at the beginning of the experiment (04.06.2010; DAY 0; 5 plants) and water was supplied to all control plants once every day (250 mL per day), whereas water supply of stressed plants was stopped. Physiological measurements and sampling of leaves took place on 07.06.2010 (DAY 3; 5 control, 5 stressed plants), 10.06.2010 (DAY 6; 5 control, 5 stressed plants) and 12.06.2010 (DAY 8; 5 control, 10 stressed plants). Due to very hot weather conditions in June 2010 the experiment was stopped after 8 days and 12 available control plants were used to restart the drought treatment with 6 control and 6 stressed plants on 11.06.2010 and all plants were measured on 15.06.2010 (DAY 5). The mean leaf temperatures at midday were: 25 ℃ (04.06.2010; DAY 0), 31.9 ℃ (07.06.2010; DAY 3), 30.8 ℃ (15.06.2010; DAY 5), 35.8 ℃ (10.06.2010; DAY 6) and 35.7 ℃ (12.06.2010; DAY 8). The mean PAR radiation per day (measured from 6:00 am till 7:00 pm) was 144.1 µmol m-2 s-1. Each plant was used only once for physiological measurements and sampling of leaves.On every day of the experiment (day 0, 3, 5, 6, 8) the pot weight and the volumetric soil moisture content (ThetaProbe ML2x and handheld data logger Moisture Meter HH2, Delta-T Devices, Cambridge, United Kingdom) was recorded. The water potential (PWSC Model 3000, Soilmoisture Equipment Corporation, Santa Barbara, USA) was determined for the 6th leaf (representing the insertion level of the shoot from the basis) of every plant and measurement day. Chlorophyll fluorescence and gas exchange parameters of light adapted leaves were determined with the 4th and 5th leaf, whereas dark adaptation was performed only with the 5th leaf. Immediately after these non-invasive measurements, the 5th leaf was harvested, frozen in liquid nitrogen and further used for the measurement of polyphenols, selected primary metabolites and volatiles (VOCs).
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor Function |
The content of different groups of primary and secondary metabolites is significantly influenced by severe drought stress in grapevine leaves. The content of the majority of the metabolites (around 60% of primary metabolites, around 85% of polyphenols and about 40% of the detected and identified VOCs) increased upon drought stress treatment. Among these especially the primary metabolites citric acid and glyceric acid were strongly influenced by the short as well as the prolonged drought stress treatment, whereas all polyphenols were only induced upon the prolonged drought stress treatment.
Click to Show/Hide
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Factor | Part | Location | NP Content | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Normal condition
|
Leaves | Vienna, Austria |
NP Content: 0.01 ± 0.00 µg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Dry 3-5 days
|
Leaves | Vienna, Austria |
NP Content: 0.01 ± 0.00 µg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
Dry 6-8 days
|
Leaves | Vienna, Austria |
NP Content: 0.02 ± 0.02 µg/g dry weight
|
||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
